3.4 Spectre des hôtes de Nosema sp. et de Mattesia sp.
Table
des matières - ![]() Précédente
- Suivante
Précédente
- Suivante![]()
La transmissibilité à d'autres espèces de coléoptères des types de spores isolés es sur P. truncatus permet de tirer certaines conclusions quant à la spécificité des hôtes des deux espèces de protozoaires considérées. Il est important de collecter des informations sur le spectre des hôtes de ces agents pathogènes dans le mesure ou ces derniers pourraient éventuellement infecter d'autres ravageurs des stocks et même, le cas échéant bloquer leur évolution.
En vue des essais de transmission, on a sélectionné, outre P. truncatus, quatre espèces d'insectes également rencontrées dans les greniers de stockage. Il s'agit d'une part d'un S. zeamais, ainsi que des ravageurs secondaires P. subdepressus, C. dimidiatus et T. castaneum. Deux bostryches, D. bifoveolatus et R. dominica ont Et! par ailleurs intégrés à cet essai du fait qu'il s'agit de ravageurs des stocks d'espèces voisines de P. truncatus et qu'ils nous ont paru à ce titre significatifs du point de vue de la transmission des protozoaires.
On trouvera au tableau 12 les résultats, détaillés pour chaque espèce de coléoptère testée, obtenus lors des essais d'inoculation des spores. Ce tableau renseigne également sur l'infection par Nosema sp. et Mattesia sp. présentée par chacun des stades de développement examinés et pour chaque espèce d'insecte, ainsi que l'infection globale (totaux et pourcentages). Les spores de Nosema et de Mattesia ayant été inoculées séparément au bostryche D. bifoveolatus, les résultats d'infection détaillés figurent également à part. En ce qui concerne l'infection globale chez les coléoptères de cette espèce, les résultats des deux inoculations ont été regroupes en un seul (cf. tabl. 12).
La profusion des résultats séparés figurent dans ce tableau permet de constater en premier lieu que toutes les espèces testées ont été contaminées par les protozoaires. A l'exception de T. castaneum, qui ne présentait pas d'infection par Mattesia, les deux agents pathogènes ont provoque une infection chez toutes les autres espèces de coléoptères concernées, ce qui exclut, aussi bien chez Nosema sp. que chez Mattesia sp., l'hypothèse d'une spécificité parasitaire au niveau du Grand Capucin du Maïs.
Tabl. 12 : Infection consécutive à l'inoculation de spores de Nosema sp. (No. sp.) et de Mattesia sp. (Ma. sp.) à 6 espèces de ravageurs des stocks
| Espèce de coleoptére |
Nombre des
individus examinés |
Nombre des individus | |||||
| par les stades de développement |
somme | par | le total | ||||
| No. sp. | Ma. sp | Do* | somme | % | |||
| S. zeamais | 195 adultes | 14 | 9 | 2 | |||
| 7 pupes | 242 | 2 | 0 | 0 | 41 | 16,9 | |
| 40 larves | 10 | 3 | 1 | ||||
| P.subdepressus | 23 adultes | 19 | 0 | 2 | |||
| 8 larves | 31 | 5 | 0 | 3 | 29 | 93,5 | |
| C.dimidiatus | 14 adultes | 8 | 1 | 2 | |||
| 3 pupes | 100 | 2 | 0 | 1 | 87 | 87,0 | |
| 83 larves | 35 | 6 | 32 | ||||
| T.castaneum | 21 adultes | 2 | 0 | 0 | |||
| 37 larves | 58 | 20 | 0 | 0 | 22 | 37,9 | |
| R. dominica D.bifoveolatus |
15 adultes | 15 | 11 | 0 | 4 | 15 | 100 |
| 68 adultes | 10 | / | / | ||||
| 21 larves | 3 | / | / | ||||
| 176 | 41 | 23 3 | |||||
| 85 adultes | / | 27 | / | ||||
| 2 larves | / | 1 | / | ||||
* Do = Double infection
L'examen des divers résultats permet de constater que, compte tenu du nombre de sujets atteints d'une double infection, la microsporidie était en règle générale beaucoup plus fréquemment représentée que la néogrégarine Dans les préparations à inoculer aux insectes, la concentration de spores de Nosema
Etait toutefois plus élevée que celle des spores de Mattesia (cf. chap 2.4.5), ce qui fait que les résultats inégaux obtenus par chacun des agents pathogènes sont vraisemblablement dus à cette différence de densité des spores au moment du traitement.
L'infection globale des diverses espèces de coléoptères a donne des résultats tout a fait hétérogènes. Alors que l on a relevé chez les ravageurs secondaires des taux d'infection allant de 38 % (sur T. castaneum) à 93,5 % (P. subdepressus), les individus de S. zeamais et de D. bifoveolatus n'étaient respectivement infestes par les protozoaires qu'à 17 et 23 %. On peut en déduire que les ravageurs secondaires des stocks étaient de toute évidence plus facilement contaminables par les protozoaires que les ravageurs des stocks aptes au forage. Notons cependant que l'on a constate une infection chez tous les représentants du bostryche R. dominica, ce coléoptère constituant en l'occurrence une exception.
Le tableau 12 permet par ailleurs de constater qu'il y avait à proportion, chez les ravageurs des stocks, davantage de larves infestées que d'adultes, ce qui vaut aussi bien pour les ravageurs secondaires que pour les ravageurs primaires. S'agissant du bostryche R. dominica, il n'a pas été possible d'établir une comparaison entre les différents stades de développement de cette espèce d'insectes vu que l'on a ici uniquement examiné des sujets adultes. Le fait que chez toutes les autres espèces de coléoptères, ce soient les larves qui aient subi la plus forte infestation semble indiquer que ce stade (tait plus propice que celui de l'imago à une infection par des protozoaires, Les résultats obtenus sur P. truncatus ayant été analogues (cf. chap. 3.3.1 et 3.3.2), il ressort de l'essai de transmission que les protozoaires considérés ont eu sur les hôtes secondaires des effets analogues à ceux observes sur l'hôte principal.
3.5 Transmission de Nosema sp. et de Mattesia sp. à Teretriosoma nigrescens
L'histeride T. nigrescens est un antagoniste de P. truncatus En tant que prédateur, il se nourrit des adultes et des larves du ravageur. La haute efficacité qu'offre 7. nigrescens dans la lutte contre l'anobie fait de cet histéridé un antagoniste de tout premier plan du ravageur des stocks (HELBIG, 1993; PÖSCHKO 1993). L'essaimage de l'insecte utile au Togo (BÖYE et al , 1992) constituait le volet central du concept de lutte biologique contre le Grand Capucin du Maïs.
A côté des nombreuses autres questions d'importance majeure sou levées par une telle démarche, il était intéressant de vérifier si les deux espèces de protozoaires décelées sur le ravageur étaient de nature à provoquer une infection chez T. nigrescens. Une affection éventuelle par ces pathogènes serait en effet susceptible d'entraver sensiblement l'évolution de l'histéridé, la mise en oeuvre future des protozoaires dans les greniers à odes paysans ayant par suite des conséquences négatives pour le prédateur. Deux essais destines à étudier le prédisposition à l'infection de T. nigrescens ont été réalisés. Dans le cadre d'une première installation expérimentale, on a placé 30 T. nigrescens adultes dans un peuplement de P. truncatus fortement infestes par des protozoaires. Lors d'un second essai, on a donne en pâture à des larves et adultes du prédateur quelques proies infectées
Concernant les résultats, il faut noter que ni les larves, ni les adultes de T. nigrescens n'ont été infectes par les protozoaires. En outre, tous les P. truncatus places dans l'installation expérimentale étant morts, on peut raisonnablement supposer que le prédateur s'était t effectivement nourri de ces proies.
Le diagnostic d'infection négatif établi au sujet de T. nigrescens corrobore les résultats de HUGER et PÖSCHKO (communication interne), qui ont examiné quelque 1000 exemplaires de l'histéridé sans déceler d'infection par les protozoaires. Il y a par conséquent lieu de supposer que les spores de Nosema et de Mattesia trouvées sur P. truncatus ne sont pas, en principe, en mesure d'infecter l'insecte utile.
3.6 Effets des insecticides associés à des spores de protozoaires en conteneur
3.6.1 Contagiosité des spores après
contamination par des insecticides (essai préliminaire)
3.6.2 Effets du pirimiphos-méthyle associé à
des spores sur la dynamique de population de Prostephanus
truncatus
En corrélation avec les insecticides habituellement utilises dans les greniers à maïs, l'application pratique de la préparation aux spores de protozoaires soulève deux questions. Il s'agit d'une part de mettre en lumière des interactions éventuelles entre les spores de protozoaires et les ruée es actives insecticides. L'application de spores dans un grenier à maïs manquerait d'efficacité si le pouvoir infectieux de ces spores était annihilé par une contamination due à des insecticides. Il était par ailleurs important de voir quels sont les effets des spores sur l'anoblie P. truncatus en cas d'application complémentaire d'une préparation chimique.
Afin de déceler d'éventuelles interactions entre les spores de protozoaires et certains insecticides, on a effectue un essai dit "préliminaire". Ce qualificatif veut dire que l'essai projeté était destine à fournir la base de l'examen ultérieur visant à déterminer les effets sur P. truncatus d'un traitement combine aux spores et aux insecticides. C'est dans le cadre d'un tel traitement combiné spores-insecticides que l'on a observe, en conteneur, la dynamique de populations du ravageur soumis à la préparation aux spores appliquée en association avec la matière active insecticide "pirimiphos-méthyle".
3.6.1 Contagiosité des spores après contamination par des insecticides (essai préliminaire)
Apres avoir stocke les spores de la préparation mixte en contact, soit avec la matière active insecticide "pirimiphos-méthyle" (Actellic), soit avec la matière active "deltaméthrine" (K-Othrine), on a teste sur des larves de P. truncatus le pouvoir infectieux des spores de Nosema et de Mattesia Les spores ayant été auparavant séparées de l'insecticide, chaque type de préparation aux spores a été inoculé à 40 larves de P. truncatus afin que l'on puisse en observer les effets infectieux sur ces larves. Une poudre aux spores conservée en état de non-contamination a servi de témoin. Les quantités de spores appliquées Etaient identiques dans toutes les variantes (cf. chap. 2.4.6).
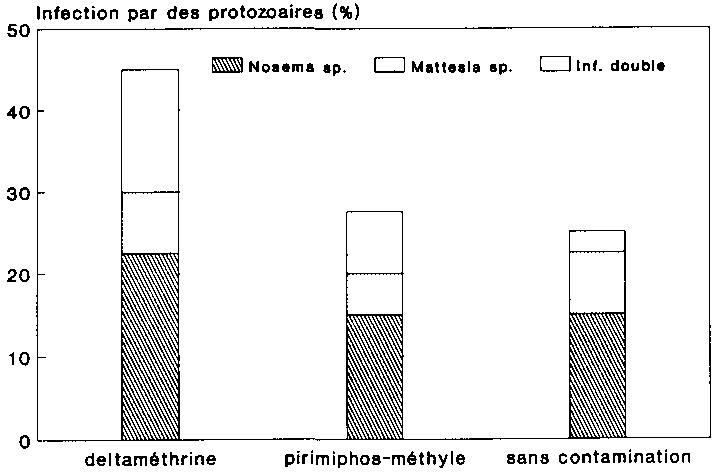
C'est l'inoculation de spores contaminées à la deltaméthrine qui, avec 45 %, a donné le taux d'infection global le plus élevé chez les larves de P. truncatus utilisées (fig. 24). Les spores contaminées à l'ester d'acide phosphorique ont, quant à elles, provoque une infection chez 27,5 % des larves, tandis que 25 % seulement des larves de l'anoblie ont été infectées à la suite d'un traitement à la poudre aux spores conservées i l'abri de la contamination par les insecticides. Le test Chi² a révélé que ces taux d'infection ne présentaient entre eux aucune différence significative.
Comme le montre par ailleurs la figure 24, les deux types de protozoaires se sont manifestes dans toutes les variantes, Compte tenu des sujets atteints d'une double infection, les larves de l'anobie étaient plus fréquemment infectées par Nosema sp. que par les espèces Mattesia. Ce résultat est conforme du fait que la préparation appliquée renfermait une quantité proportionnellement supérieure de spores de Nosema (cf. Chap. 2.4.6). On ne peut donc pas en conclure que les deux types de spores différent au niveau du pouvoir infectieux après contamination par les insecticides considérés.
L'infection subie par les larves permet de conclure en résume que la contamination par les matières actives insecticides n'a pas amoindri la contagiosité des deux types de spores. Le taux d'infection relativement élevé des deux agents pathogènes constaté sur les larves à la suite du traitement aux spores contaminées par de la deltaméthrine permet même de supposer que le stockage commun avec l'insecticide "K-Othrine" stimule le pouvoir infectieux des spores.
L'essai décrit dans la suite avait pour but de déterminer les effets, sur la population de P. truncatus, de traitements aux spores de protozoaires associées au pirimiphos-méthyle, une matière active insecticide. Nous allons brièvement indiquer ici les raisons qui ont préside du choix d'une liaison ester d'acide phosphorique pour une mise en oeuvre combinée à des spores de protozoaires.
On sait par les ouvrages spécialisés que la matière active "pirimiphos-méthyle" n'est pas en mesure de paralyser durablement le développement de P. truncatus (GOLOB et a 1, 1985). Cette efficacité insuffisante de l'ester d'acide phosphorique nous a conduits à ver fier si l'association du principe actif a des spores de protozoaires était de nature à en renforcer l'efficacité. L'idée de départ était que la contamination de l'insecte par les protozoaires pouvait éventuellement le sensibiliser davantage au principe actif chimique.
On a pris en l'ocurrence des mâles et femelles sains du ravageur des stocks, que l'on a pincés dans des récipients à raison de 10 sujets par conteneur. Ces animaux ont reçu respectivement un traitement unique à la préparation aux spores, au principe actif "pirimiphos-méthyle, ou encore ils ont été traités du moyen de préparations combinées de poudre aux spores et de pirimiphos-méthyle. Les doses employées lors de l'application de l'insecticide seul et des traitements combinés étalent respectivement de 0.01 et de 0.005 mg de pirimiphos-méthyle par g de substrat (cf. tabl. 3).
Il a été procédé à répétitions par variante. A l'issue d'une période de 4, 8 et 12 semaines, les individus ont été dénombrés dans chacune des variantes. Dans les peuplements traites aux spores ou aux préparations combinées, on a prélevé à l'issue de chacune des périodes indiquées un échantillonnage de 50 animaux par stade de développement et procédé sur eux à une recherche d'infection par les protozoaires.
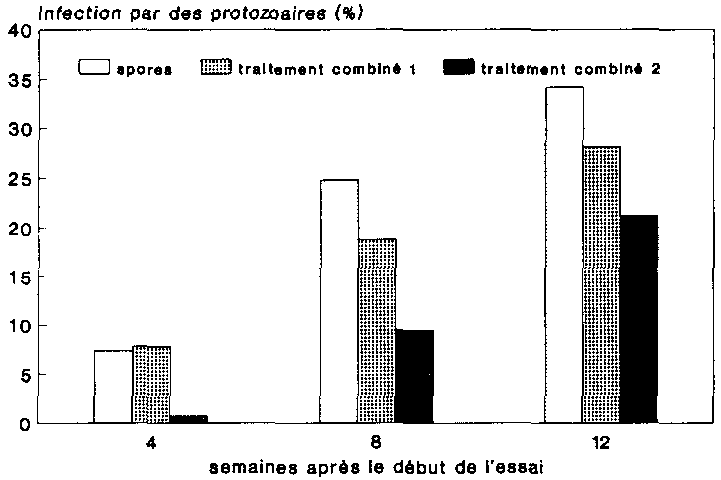
Si l'on compare le développement de P. truncatus sous l'effet des divers traitements, on constate que ce sont les populations non traitées qui ont connu le développement la plus favorable (fig. 25, témoin). On a pu observer pendant toute la durée de l'essai une croissance de ces populations, qui comptaient au bout de 12 semaines 2390 animaux en moyenne. Les populations de ravageurs traitées à la préparation aux spores ont connu un développement plus mauvaise que le témoin (fig. 25, spores). Avec quelque 1500 sujets au bout de 12 semaines, les animaux étaient en l occurrence nettement moins nombreux qu'au sein de la population non traitée.
Sous l'effet du pirimiphos-méthyle, le nombre de sujets à se développer a été dans tous les cas inférieur à ce qu'il a été sur le témoin et à la suite de l'application de spores. Si le traitement à 0,005 mg de pirimiphos-méthyle par g de substrat (fig. 25, Actellic 1) a permis d'observer pendant toute la durée de l'essai une augmentation du nombre d'individus, on a constate en revanche après 12 semaines une réduction moyenne des populations à 612 animaux seulement. Les populations traitées à 0,01 mg de pirimiphos-méthyle par g de substrat se sont reproduites dans l'ensemble encore plus mal (fig. 25, Actellic 2), puisqu'elles ne comptaient plus en moyenne que 114 individus au bout de 12 semaines.
Dans les conteneurs traites aux préparations combinées, la dynamique de la population s'est maintenue pendant toute la durée de l'essai à un niveau supérieur à ce qu'elle était chez les P. truncatus traites à l'Actellic (cf. fig. 25). Ayant déjà atteint une densité relativement élevée au bout de 4 semaines d'essai, les populations ont été en mesure de continuer à se multiplier également après 8 semaines' et cela pour les deux traitements combines. Au bout de 12 semaines, on a dénombré en moyenne dans la variante traitée aux spores additionnées de 0,005 mg de pirimiphos-méthyle, par g de substrat (traitement combiné 1) 500 animaux. On a constaté ici un léger recul du nombre de sujets par rapport à ce qu'il était au bout de 8 semaines. Les populations traitées à 0,01 mg de principe actif par g de substrat additionné de spores (traitement combiné 2) comptaient au bout de 12 semaines 367 individus, soit à peu près le même nombre qu'au bout de 8 semaines.
Le taux de mortalité des populations d'anobies ayant subi les divers traitements a été établi à chaque fois pour tous les individus d'un même conteneur. Le tableau 13 indique les taux de mortalité moyens par variante à chacune des dates dévaluation. Ce sont les populations traitées à l'Actellic hautement concentré (Actellic 2) qui présentaient le taux de mortalité le plus élevé. Au bout de 8 semaines, on dénombrait en effet ici 70,1 % dé sujets morts et 65 4 % au bout de 12 semaines. Dans toutes les autres variantes en revanche, le nombre de coléoptères morts atteignait tout au plus 33,6 %, ce qui se situe nettement en deçà des taux de mortalité des populations traitées à l'Actellic 2. On notera en l'occurrence la mentalité initiale élevée observée au sein des populations de ravageurs traitées uniquement a l'insecticide. Au bout de quatre semaines, le taux de mortalité s'établissait respectivement à 30 % (Actellic 1), et à 36 % (Actellic 2), alors que les populations des autres variantes accusaient des taux de mortalité allant de 1 à 6 % seulement.
Tabl 13: Mortalité chez Prostephanus truncatus à la suite de divers traitements à la préparation aux pores et à l'Actellic après 4,8 et 12 semaines a essai
| Variantes | Mortalité (%) |
||
| 4 | 8 | 12 | |
| temoin | 3,3 | 10,5 | 27,1 |
| spores | 1,3 | 21,3 | 33,6 |
| Actellic 1 | 29,9 | 29 ,8 | 21,0 |
| Actellic 2 | 36,3 | 70,1 | 65,4 |
| traitement | 3,4 | 22,6 | 33,5 |
| combinés 1 | |||
| traitement | 6,2 | 11,3 | 17,6 |
| combinés 2 | |||
* variantes: pour les abréviations, se reporter à la fig. 25.
On trouvera sur la figure 26 les résultats de l'infection par les protozoaires chez les populations traitées aux spores (spores, traitements combinés 1 et 2). En ce qui concerne les taux d'infection, il s'agit de l'infection globale moyenne pour tous les stades de développement à une date donnée, 50 sujets ayant été examines à chaque fois.
On a observe pour toutes les variantes une augmentation des taux d'infection à chaque date d'évaluation. A la différence des taux d'infection obtenus à la suite du traitement simple aux spores, les traitements combines se sont traduits par des taux d'infection moindres au niveau des individus. On notera ici avec intérêt que le traitement combiné comportant la concentration de pirimiphos-méthyle plus faible (traitement combiné 1) a provoque au sein des populations un taux d'infection proportionnellement supérieur à celui résultant du traitement qui faisait intervenir une concentration de matière active plus élevée (traitement combiné 2).
Les différences constatées d'une variante a l'autre indiquent clairement que l'application de spores seules a permis aux protozoaires de se propager plus facilement dans la population de l'Hôte que lorsqu'elles ont été appliquées en association avec un traitement a l'Actellic. Il semble que la concentration de matière active insecticide introduite dans le conteneur ne soit pas sans influence sur la propagation de l'agent pathogène, puisque aussi bien les traitements hautement concentres ont été suivis d'un taux d'infection à proportion plus faible que ceux qui contenaient une moindre concentration de matière active.
C'est à l'issue de 12 semaines d'essai que l'on a déterminé chez les populations traitées aux spores le taux d'infection le plus élevé, c'est-à-dire 34 %. Les autres taux d'infections étant demeures inférieurs à 30 %, l'infection s'est donc avérée faible dans l'ensemble. Il n'est pas possible, du moins pour l'instant, de tirer des conclusions quant à ce bas niveau d'infection vu que les doses de spores inoculées étaient relativement élevées. Il est toutefois concevable que la préparation inoculée n'ait été que faiblement infectieuse.
En résumé, on peut dire que les traitements aux spores de protozoaires et ceux à la matière active pirimiphos-méthyle ont eu des effets réducteurs sur la dynamique des populations de P. truncatus. L'application des variantes aux insecticides s'est traduite par des taux de mortalité relativement élevés qui n'ont été constatés ni pour les traitements combines, ni sous le seul effet des spores. Comparées aux populations traitées à l'Actellic, celles qui avaient été traitées au moyen des préparations combinées ont produit dans l'ensemble davantage de sujets. Ces résultats permettent de conclure que le mélange de poudre insecticide et de poudre aux spores donne vraisemblablement naissance à des interactions antagoniques entre la matière active insecticide et les spores de protozoaires. Cette hypothèse expliquerait d'une part le baisse d'efficacité de l'insecticide contre le ravageur, et d'autre part les faibles taux d'infection sur P. truncatus dans les variantes à préparations combinées.
Ajoutons pour terminer qu'il n'a pas été possible de tirer de conclusions quant aux effets de l'ester d'acide phosphorique sur des P. truncatus infectes par des protozoaires en raison du faible niveau d'infection constaté dans l'ensemble.
3.7 Mise en évidence de spores de protozoaires dans des produits résiduels du stockage du maïs
Du point de vue de la mise en oeuvre pratique des spores de protozoaires il serait souhaitable, et cela pour des raisons évidentes de réduction des coûts, que les paysans se chargent eux-mêmes de confectionner la préparation aux spores de protozoaires. Nous allons examiner dans la suite les possibilités dont dispose l'utilisateur pour fabriquer cette préparation de manière autonome.
Au cours du processus d'extraction des grains consécutif du stockage du maïs en épis tel qu'il est traditionnellement pratique dans le sud du Togo, il se forme des résidus n'ayant aucune valeur nutritive. Il s'agit en l'occurrence de spathes, de rachis, de brisures de grains, d'ivraie, et, en cas d'infestation de l'entrepôt par des ravageurs' d'insectes parasites et de la poussière de forage qu'ils produisent. Afin d'étudier les possibilités de fabrication d'une poudre aux spores à partir des insectes existants, on a collecte les résidus d'un stockage de maïs, que l'on a ensuite examinés à la recherche de spores éventuelles.
Durant une période de cinq mois, on a prélevé chaque mois dans un grenier à maïs 50 épis Les résidus provenant de l'extraction des grains ont été stockes dans un fut pratiquement hermétique. Apres avoir opéré le dernier prélèvement d'épis dans le grenier, les déchets conservés ont été triés, et les éléments grossiers, à savoir les spathes, les axes des épis et les brisures de grains, ont été séparés des résidus fins. A partir du mélange résiduel, constitué de poussière de farine, de coléoptères et d'ivraie, on a préparé une poudre fine, dont on a prélevé trois échantillons de 20 mg chacun. Ces échantillons de poudre ont été ensuite examinés selon la méthode exposée plus haut afin de déterminer la densité des spores (cf. chap. 2.3.5).
L'analyse des échantillons de farine a révélé une concentration moyenne de 7,5x104 de spores de Mattesia et de 5x105 de spores de Nosema dans I ml d'eau distillée. Comparées à celles des préparations utilisées lors des essais en conteneurs, ces valeurs étaient inférieures de 1 à 2 puissances de dix. On a donc mis en évidence dans la poudre obtenue à partir des résidus de stockage du maïs une concentration de spores très élevée. Les spores de protozoaires détectées dans la farine proviennent vraisemblablement des cadavres d'insectes contenus dans le mélange pulvérisé.
3.8 Application de la préparation aux spores de protozoaires dans des conditions semi-pratiques
3.8.1 Mise en oeuvre de la préparation aux
spores dans un grenier à maïs traditionnel-essai I (1990) à
Cacaveli
3.8.2 Mise en oeuvre de la préparation aux
spores dans un grenier à maïs traditionnel - essai II (1990/91)
à Davié
Après examen des résultats des essais réalisés en conteneurs, nous allons décrire dans la suite ceux qui avaient pour but de tester, dans des conditions proches de la réalité, les effets de la préparation aux spores. Deux essais similaires de stockage, réalisés selon les méthodes traditionnelles. de stockage du maïs en usage dans cette région, se sont déroulés dans le sud du Togo. Dans le cadre de l'essai de stockage I, le maïs a été entrepose de février à juillet 1990 (pendant la "petite saison de stockage"), tandis que l'essai II avait lieu d'août 1990 à janvier 1991 (durant la "grande saison").
Table
des matières - ![]() Précédente
- Suivante
Précédente
- Suivante![]()

{kind=link}
{kind=link}
{kind=link}