Table
des matières - ![]() Précédente
- Suivante
Précédente
- Suivante![]()
3.1 Présence et distribution des protozoaires
sur Prostephanus truncatus au Togo
3.2 Description et classification des espèces
de protozoaires
3.3 Pathogénicité en laboratoire des
protozoaires affectant Prostephanus truncatus
3.4 Spectre des hôtes de Nosema sp. et de
Mattesia sp.
3.5 Transmission de Nosema sp. et de Mattesia
sp. à Teretriosoma nigrescens
3.6 Effets des insecticides associés à des
spores de protozoaires en conteneur
3.7 Mise en évidence de spores de protozoaires
dans des produits résiduels du stockage du maïs
3.8 Application de la préparation aux spores de
protozoaires dans des conditions semi-pratiques
3.1 Présence et distribution des protozoaires sur Prostephanus truncatus au Togo
La région méridionale du Topo constitue la principale zone de distribution du ravageur P. truncatus dans ce pays. Le maïs, qui y fait l'objet de deux récoltes annuelles, est traditionnellement stocké en épis, avec les spathes, lors de la "grande saison de stockage" (juin/juillet) et de la "petite saison de stockage" (décembre/janvier). Les greniers à maïs, qui sont construits en bois, offrent à I'anobie de bonnes chances de subsistance entre deux périodes de stockage puisque aussi bien le ravageur est en mesure de pénétrer dans le bois (DETMERS, 1988). Le recensement de la présence de P. truncatus dans les greniers de stockage a eu lieu en janvier et février de l'année 1989, à une époque où une partie de la récolte de maïs précédente avait déjà été prélevée et où l'on venait seulement de rentrer la nouvelle récolte, ce qui fait que l'anobie n'a pu être découverte que dans des résidus de stockage anciens.
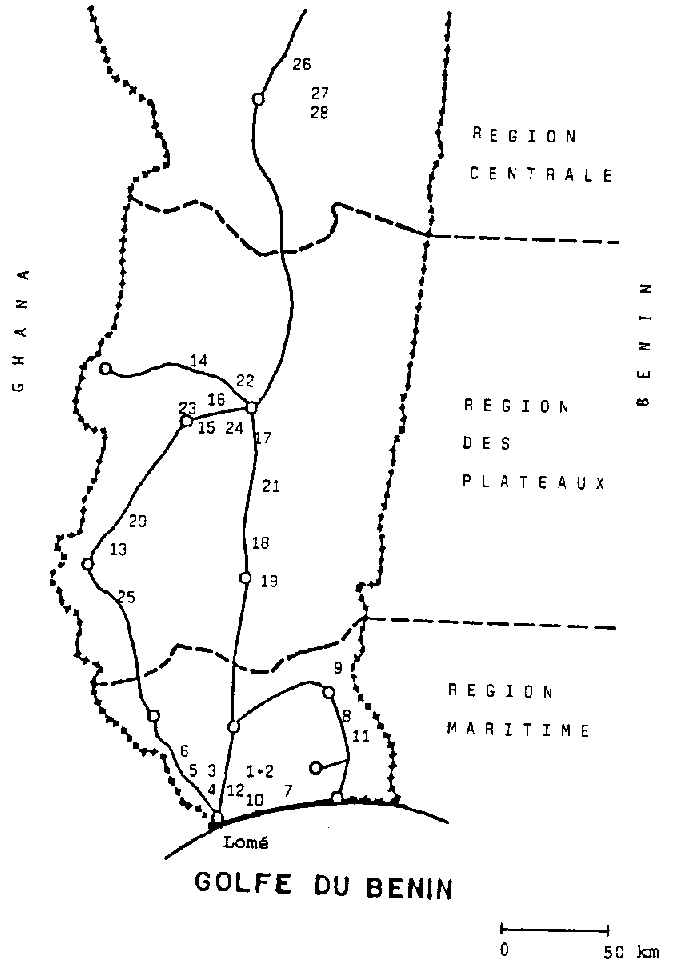
La figure à représente la zone dans laquelle s'est déroulée l'enquête. Les chiffres indiquent les lieux de découvert de P. truncatus dans la "Région Maritime", la "Région des Plateaux" et la "Région Centrale". Sur les 103 villages visites, 28 présentaient dans leurs greniers à maïs une infestation par P. truncatus. On trouvera au tableau 5 les noms des villages, ainsi que les résultats des examens de détection des agents pathogènes sur le ravageur.
On a décelé sur les anobies examinées deux espèces de protozoaires. Il s'agit tout d'abord d'une néogrégarine. Un protozoaire de ce type ayant déjà été découvert au Togo sur P. truncatus dans le cadre d'un essai similaire (LELIVELDT et al., 1988), nous allons examiner dans la suite la question de savoir si les deux agents pathogènes sont identiques. L'autre protozoaire, qui fait partie des microsporidies, a été décelé au Topo pour la première fois. Son identité fera également dans la suite l'objet d'une étude plus approfondie. Les deux espèces de protozoaires à analyser sont provisoirement désignées au tableau 5 sous le nom de "néogrégarine" et de "microsporidie".
Alors que la néogrégarine était présente dans les trois régions considérées, l'espèce de microsporidie n'a pas a été détectée dans la région centrale, mais uniquement dans les régions méridionales. La néogrégarine était dans l'ensemble plus répandue. Elle s'est manifestée en outre plus fréquemment que l'autre espèce, puisque l'on a pu mettre en évidence sur 14 sites des coléoptères infestes par la néogrégarine, tandis que la microsporidie n'a été trouvée sur l'anobie que dans 7 localités. Deux P. truncatus découverts à Agoenyivé ("Région Maritime") présentaient la particularité d'être infestés simultanément par les deux agents pathogènes (cf. tabl. 5).
De manière générale, l'infection naturelle du ravageur des stocks par les protozoaires peut être considérée comme faible, puisque aussi bien dans la majorité des cas 7 animaux sur 24 étaient infestés. Avec 24 exemplaires par site de découverte, le nombre de coléoptères examines est demeure très limité, dans la mesure où il s'agissait seulement de réaliser une mise en évidence qualitative du spectre des agents pathogènes naturels de P. truncatus Les deux espèces de protozoaires découvertes ont été isolées. à la fois aux fins d'identification et en vue des essais d'inoculation prévus.
Tabl. 5 : Mise en évidence de protozoaires sur des Prostephanus truncatus originaires de trois régions infestées du Togo
| Villages
Région infestés ravageurs |
Infection des | ||||||
| vivants | ravageurs | morts | |||||
| Maritime | |||||||
| 1 | Adyougba | 0 | (12)* | 0 | ( 9) | ||
| 2 | Aveime | 0 | (12) | 0 | ( 9) | ||
| 3 | Logopé | 0 | (12) | 0 | (12) | ||
| 4 | Gbonvé | 2 Ne** | (12) | 2 Ne | (12) | ||
| 5 | Sanguera | 0 | (12) | 7 Ne | (12) | ||
| 6 | Aképé | 0 | (12) | 1 Ne | (12) | ||
| 7 | Lébé | 3 Ne | (12) | 3 Ne | (12) | ||
| 8 | Logomé | 1 Ne | (12) | 1 Ne | ( 7) | ||
| 9 | Gboto- Assigamé |
0 | (12) | 1 Ne | (12) | ||
| 10 | Kélégougan | 3 Mi** | (12) | 2 Mi | ( 5) | ||
| 11 | Atitogan | 1 Ne | (12) | 1 Mi | (12) | ||
| 12 | Agoenyivé | 0 | (12) | 2 Ne + | |||
| 2 Mixte** | (12) | ||||||
| Des Plateaux | |||||||
| 13 | Kpime-Tomegbe | 0 | (12) | 0 | (12) | ||
| 14 | Oudje | 0 | (12) | 0 | (12) | ||
| 15 | Ezimé | 0 | (12) | 0 | (12) | ||
| 16 | Ayomé | 0 | (12) | 0 | ( 4) | ||
| 17 | Avété | 0 | (12) | 0 | (12) | ||
| 18 | Agbatitoe | 0 | (12) | - | |||
| 19 | Notse | 0 | (12) | 0 | (12) | ||
| 20 | Levié Apodomé |
1 Ne | (12) | 1 Ne | (12) | ||
| 21 | Glei | 0 | (12) | 1 Ne | (12) | ||
| 22 | Evoubete-Agbede | 0 | (12) | 1 Mi | (12) | ||
| 23 | Adjahun | 1Mi(12) | 0 | (12) | |||
| 24 | Koutoukpa | 0(12) | 1 Mi | (12) | |||
| 25 | Touvé Agbessia |
3 Mi | (12) | 4 Ne | (12) | ||
| Centrale | |||||||
| 26 | Labidé | 0 | (12) | 1 Ne | (12) | ||
| 27 | Niamgoulam | 0 | (12) | 3 Ne | (12) | ||
| 28 | Atara | 1 Ne | (12) | 1 Ne | (12) | ||
* entre () = Nombre de coléoptères examines
** Ne = Infection par la néogrégarine,
Mi = Infection par la microsporidie
Mixte = Infection par les deux espèces de protozoaires
3.2 Description et classification des espèces de protozoaires
3.2.1 Morphologie des spores
3.2.2 Cycle évolutif de Nosema sp,
3.2.3 Histopathologie de Nosema sp.
Les deux types de protozoaires mis en évidence sur P. truncatus ont été soumis a un examen plus approfondi. Les figures 4 et 5 permettent de reconnaître les spores de ces deux types de protozoaires A l'état naturel. Les deux types considérés diffèrent fortement du point de vue de la forme et de la taille. Alors que la spore des microsporidies est plutôt petite et possède une forme ovale a ovoïde (fig. 4), les spores de la néogrégarine affectent la forme d'un citron et sont beaucoup plus grandes (fig. 5).
La forme caractéristique de la spore des microsporidies et la présence d'un filament polaire à l'intérieur permettent de conclure avec un haut degré de certitude qu'il s'agit d'une microsporidie du genre Nosema (POINAR & THOMAS, 1978). La morphologie des spores fournissant un critère de distinction entre les espèces Nosema on a par conséquent détermine ici les dimensions, la longueur des filaments polaires et le rapport nucléaire à l'intérieur des spores, puis compare ces caractéristiques à celles de trois autres espèces Nosema ainsi qu'à trois isolats de spores.
En ce qui concerne les espèces qui nous occupent, il s'agit d'une part de N. whitei sur ses deux hôtes principaux, T. confusum et 7. castaneum (LIPA, 1968: MILNER, 1972d), ainsi que de N. weiseri sur R. dominica (LIPA, 1963) et N. oryzaephili sur Oryzeaphilus surinamensis (BURGES et al., 1971). Enfin, on a inclus dans la comparaison trois isolais de spores de diverses microsporidies trouvées par PURRINI (communication interne) sur P. truncatus en Tanzanie.
Les spores de Nosema examinées ici mesuraient en moyenne 3,3 mm de long, les mesures effectuées ayant révélé par ailleurs Une largeur moyenne de 2,1 µm. Ces données correspondaient assez bien a celles concernant les spores de N. weiseri, alors que N. whitei et N. oryzaephili possèdent des spores de dimensions très supérieures. S'agissant de la comparaison des spores examinées avec les trois isolats trouvés en Tanzanie, il s'est avéré que si les dimensions des spores de l'isolat I étaient assez similaires a celles résultant de nos propres mesures, les isolats 2 et 3 présentaient des spores beaucoup plus grandes que celles mesurées chez Nosema sp..
Autre caractéristique permettant la distinction entre les spores de Nosema; la longueur des filaments polaires. Concernant cette caractéristique chez les trois isolats de spores de P. truncatus, on trouve uniquement chez PURRINI (communication interne) des indications relatives à la forme de spirale du filament contenu dans les spores. La comparaison se limite donc aux spores de Nosema décrites en corrélation avec les autres ravageurs des stocks. Les mesures auxquelles nous avons procédé sur 20 filaments polaires ont permis d établir une longueur moyenne de 117,6 µm. Cette valeur se rapprochait le plus de la longueur du filament polaire des spores de N. whitei, qui est de 112 µm sur T. castaneum (MILNER, 1972a).
PURRINI (communication interne) n'indiquait le rapport nucléaire des spores que pour 2 isolats (1. 1 et 1.2). Chez LIPA (1968), cette indication fait totalement défaut au sujet de N. weiseri. Les spores de Nosema de nos propres examens possédaient deux noyaux par spores. A l'exception de l'isolat de spores 2, qui provenait de Tanzanie et ne présentait qu'un seul noyau, on a également trouvé dans les spores de l'isolat tanzanien 1, de même que dans celles de N. oryzaephili et de N. whitei, deux royaux.
Les dimensions des spores de la néogrégarine, telles qu'elles ont été constatées, figurent au tableau 6. Les résultats de ces mesures de longueur et de largeur concordent bien avec les indications fournies par LIPA & WOHLGEMUTH (1986), et LEIVELDT (1990), qui décrivent une espèce Mattesia trouvée au Topo sur le ravageur P. truncatus. On peut par conséquent postuler qu'il s'agit de la même espèce. Cette hypothèse a été renforcée par les stades intermédiaires de la néogrégarine observas au cours de nos propres examens, puisque aussi bien ces stades sont conformes a la description donnée par LELIVELDT (1990) du cycle évolutif d'espèce Mattesia.
Tabl. 6 : Dimensions des spores de Mattesia trouvées au Togo sur des Prostephanus truncatus (l'état naturel)
| En µm | D'aprés LIPA et al. (1986) |
D'aprés
LELIVELDT (1990) |
D'aprés ses propre mésurages |
| Longueur | 11,6-14 | 12,6 | 10,6-12,5 |
| Largeur | 6,7- 7,8 | 7,2 | 6,3-7,8 |
3.2.2 Cycle évolutif de Nosema sp.
Les cycles évolutifs des microsporidies renseignent sur le mode de multiplication du protozoaire à l'intérieur de son hôte. De manière générale, on peut distinguer une multiplication sexuée et une multiplication asexuée, qui permettent à l'agent pathogène de se propager et de se reproduire dans les cellules de l'hôte (WEISER, 1961 LIPA, 1974). On désigne sous le terme de schizogonie l'évolution asexuée. Les schizontes (appelés également mérontes) se multiplient par deux divisions consécutives, résultant dans une colonisation intense des tissus de l'hôte (cf. fig. 6). La reproduction sexuée commence avec la formation de diplokaryen, chez lesquelles les deux noyaux des schizontes fusionnent. C'est à partir de ces stades que se développent les sporontes, qui se transforment ensuite en sporoblastes. Ceux-ci constituent les stades précurseurs de la jeune spore. Avec l'apparition de la forme mure de la spore, l'évolution est parvenue à son terme et un nouveau cycle peut commencer. En tant que stade permanent des microsporidies, les spores sont très résistantes et peuvent souvent demeurer longtemps viables dans le sol ou dans le corps d'insectes morts. Les spores assurent donc la survie du protiste et possèdent la capacité d'infecter à nouveau des insectes lorsqu'elles sont ingérées.
Caractéristique des microsporidies, le cycle évolutif décrit ne diffère pas fondamentalement chez la plupart des espèces Nosema. Ajouté 4 d'autres caractéristiques, comme la morphologie des spores et les affinités organiques à l'intérieur de l'hôte, il peut cependant fournir de précieuses indications pour l'identification du protozoaire, ce qui explique que l'on ait mis ici en lumière le cycle évolutif de l'espèce Nosema considérée afin de faciliter la suite du processus d'identification.
On a observe dans un premier temps l'apparition de schizontes à structure cytoplasmique très dense, mononucléés ou binuclées, plus rarement quadrinucléés (fig. 6 a). Ces schizontes se reproduisaient par division. C'est à partir d'eux que se sont développes les sporontes, mononuclées ou binuclées, au plasma plus clair (fig. 6 b). La division de chaque sporonte (fig. 6 c) a donne naissance à deux sporoblastes (fig. 6 d), lesquels se sont transformés en spores après différents stades précurseurs. Les deux noyaux colores sont bien visibles dans les jeunes spores (fig. 6 e).
Au vu du cycle évolutif tout à fait caractéristique présente par le protozoaire, ainsi que des particularités de ses spores décrites au chapitre précédent, l'espèce de microsporidie a laquelle nous avons affaire ici appartient à la famille des Nosematidae et au genre Nosema NÄGELI (d'après POINAR & THOMAS, 1978).
Le cycle de cette Nosema a été comparé a ceux des espèces N. whitei, N. weiseri et N. oryzaephili, qui ont été décrits dans la littérature spécialisée. PURRINI (communication interne) s'étant limité a la mise en évidence sur P. truncatus des spores mûres de l' "isolat 1", il n'a pas été possible d'établir de comparaison en ce qui concerne l'évolution. BURGES et al. (1971) ont observé dans le cycle de N. oryzaephili une division de chacun des sporontes en deux sporoblastes. Dans le cycle évolutif représenté ici, le sporonte se divisait également en deux sporoblastes, donnant chacun naissance à une spore (cf. fig. 6 c et d).
LIPA (1968) a décrit en revanche dans le cycle de N. weiseri la formation des sporoblastes a partir d'un sporonte unique. On n'a donc pas constaté ici de division. En ce qui concerne l'évolution de l'espèce N. whitei, MILNER (1972a) et LIPA (1968) n'ont observe là encore aucune division des sporontes au cours du cycle évolutif.
3.2.3 Histopathologie de Nosema sp.
L'examen histopathologique de coupes sagittales colorées des larves de P. truncatus infestées par Nosema a révélé une infection du niveau de plusieurs organes. Le figure 7 montre les principaux foyers infectieux du protozoaire. La microsporidie infectait les tissus musculaires avec une certaine virulence, Outre quelques parties de la musculature du corps, la musculature intestinale présentait des foyers d'infection avancés. La chaîne ganglionnaire ventrale, de même que l'organe dorsal, (talent également infestes par l'agent pathogène. La matrice trachéenne et l'épiderme étaient eux aussi nettement infestés, bien qu'à un moindre degré. On n'a pas décelé d'infection dans les séries de coupes pratiquées du niveau de l'épithélium intestinal. Il est par ailleurs intéressant de noter que l'on a observe des colonies très denses de l'agent pathogène dans de larges secteurs du corps adipeux (cf. fig. 7).
Les résultats obtenus permettent de conclure à une nette affinité de l'agent pathogène envers les tissus adipeux de l'animal hôte. Cette propriété est cependant commune A de nombreuses microsporidies pathogènes des insectes (WEISER, 1961).
Au vu de la comparaison de la morphologie des spores, du cycle évolutif et de l'histopathologie, nous constaterons pour terminer que l'espèce Nosema découverte au Togo pourrait être identique à N. oryzaephili. En outre, eu égard à la forte similarité entre les dimensions des spores isolées en Tanzanie et celles des spores découvertes sur P. truncatus au Togo, il s'agit éventuellement des mêmes. Il n'est toutefois pas encore possible de classifier la microsporidie avec certitude, du fait que certains critères essentiels, comme la comparaison des dimensions aux autres stades d'évolution, n'ont pas encore été établis.
3.3 Pathogénicité en laboratoire des protozoaires affectant Prostephanus truncatus
3.3.1 Effets pathogènes sur des larves de
Prostephanus trunchatus
3.3.2 Effets pathogènes sur des adultes de
Prostephanus truncatus
3.3.3 Incidences d'une infection par Mattesia
sur la fécondité et la longévité de Prostephanus truncatus
3.3.4 Mise en évidence de spores dans des
excréments de Prostephanus truncatus infestés
3.3.5 influence des spores de protozoaires sur
la dynamique des populations de Prostephanus truncatus
3.3.6 Contagiosité des spores de protozoaires
après stockage
La condition fondamentale à la mise en oeuvre pratique des agents pathogènes est qu'ils présentent. Un haut degré de pathogénicité vis-à-vis de l'organisme cible. Dans le but de déterminer les effets pathogènes de Nosema sp. et de Mattesia. sp. sur P. truncatus, on a entrepris toute une série d'essais d'inoculation en conteneur.
Dans un premier temps, on a inoculé les deux types de spores aux différents stades larvaires, ainsi qu'aux adultes du ravageur, et observé la réaction de ces animaux à un traitement c l'un ou l'autre de ces protozoaires. Dans le cadre d'un essai par couples, on a par ailleurs étudié l'influence de l'infection par Mattesia sur la fécondité et la longévité de P. truncatus
Concernant les possibilités de contamination du protozoaire au sein de la population par évacuation des spores, il a été procédé a une analyse des matières fécales d'animaux atteints.
Outre les examens effectués sur des individus, on a également observé l'évolution de la population de ravageurs à la suite d'un traitement aux spores. Dans le cadre d'un essai, on a place en observation des P. truncatus infestes naturellement et, dans un essai parallèle, des coléoptères sains, et note leur évolution sur plusieurs semaines. Désirant élucider un aspect important de l'application pratique des spores, on a testé leur capacité de résistance par deux essais d'inoculation de spores, lesquelles avaient été soumises auparavant à un stockage plus ou moins prolongé.
3.3.1 Effets pathogènes sur des larves de Prostephanus truncatus
Nosema sp.
Dans le but de déterminer la pathogénicité de Nosema sp., on a inocule des spores de Nosema aux trois stades larvaires de P. truncatus Au bout de 19 jours (pour les L3), de 26 jours (pour les L2) et de 33 jours (pour les L1), on a note l'évolution et établi le taux de mortalité des larves utilisées, ainsi que le nombre d'animaux infestes
La haute mortalité initiale constatée au cours de l'essai 1, aussi bien chez les larves traitées que chez les larves non traitées, indique à d'évidence que cet essai s'était déroulé dans des conditions défavorables. On y avait utilise de la farine de maïs non tamisée, de sorte que les particules de farine grossières avaient probablement cause la mort de nombreuses larves. Dans leur environnement naturel, les larves se développent en effet dans la farine plus fine produite par leurs congénères au cours de leur activité de forage. On a donc procédé à un second essai, en employant cette fois de la farine finement tamisée.
Au vu des deux essais réalisés, on peut conclure que c'est le premier stade larvaire de l'anobie qui présente le plus haut degré de prédisposition envers les spores de Nosema Alors que les larves du stade L2 montraient {gaiement une haute disposition à l'infection par Nosema on a établi pour le stade L3 un taux d'infection très inférieur. Seul le stade L2 présentait à la suite du traitement aux spores un taux de mortalité relativement élevé. Chez les deux autres stades larvaires, l'infestation n'a pas entraîné d'augmentation du taux de mortalité.
Mattesia sp.
S'agissant de l'inoculation de spores de Mattesia à des larves, on a constaté qu'au cours de l'un des deux essais seulement l'inoculation avait provoque chez les L1 et L2 de P. truncatus une infestation. A l'instar des résultats obtenus sur l'espèce Nosema les L3 ont réagi également plus faiblement au traitement a base de spores de la néogrégarine. Une influence de l'agent pathogène sur la mortalité n'a pu être mise en évidence que pour le stade L1. Les L1 et L2 ont réagi par ailleurs à l'infection par Mattesia par une inhibition de la croissance.
3.3.2 Effets pathogènes sur des adultes de Prostephanus truncatus
Nosema sp.
Afin d'établir la prédisposition des adultes du ravageur des stocks, on leur a inocule des spores de Nosema. Deux concentrations différentes ont été appliquées lors des traitements. Les résultats d'infestation sur les imagos figurent au tableau 7. Seule la préparation de poudre aux spores la plus concentrée a provoqué une infection chez les coléoptères. On n'a cependant pas constaté d'effets mortels de l'agent pathogène.
Tabl. 7 : Nombre d'adultes de Prostephanus truncatus infectés aprés inoculation de deux concentrations différentes de spores de Nosema
Concentration |
Adultes |
|
| vivants | morts | |
| 8 x 106 | 76 (8) | 4 (1) |
| 8 x 105 | 77 (0) | 3 (0) |
| Temoin | 78 | 2 |
* entre ( ) = nombre de coléoptères infestés
Mattesia sp.
On a également inocule aux coléoptères de P. truncatus deux concentrations différentes de spores de Mattesia sp. Le tableau 8 indique le nombre de sujets infectes à l'issue d'une période d'incubation de trois semaines. Le nombre de coléoptères infestés par l'agent pathogène était très restreint. Sur les 120 adultes objets de l'essai, seuls 4 individus dans un cas, et à dans l'autre, étaient infectés. La plupart des insectes infectes ayant survécu, on ne peut pas, par conséquent, parler 13 non plus d'effets mortels sur les imagos.
Tabl. 8 : Nombre d'adultes de Prostephanus truncatus
infectes après inoculation de spores de Mattesia
| Concentration
des spores par g substrat |
Adultes |
|
| vivants | morts | |
| 4 x 104 | 109 (2)* | 11 (2) |
| 2 x 104 | 116 (3) | 4 (0) |
| Temoin | 110 | 10 |
* entre () = nombre de coléoptères infectes
Les effets exerces sur la reproduction de l'animal hôte constituent un important critère d'appréciation des propriétés pathogènes d'un protozoaire. Dans le cadre de l'essai présenté ci-après, on a observé dans chaque groupe dix couples de P. truncatus Le premier rassemblait uniquement des individus sains (variante 1), le second des mâles (variante 2) et le troisième des femelles (variante 3) infestés par l'espace Mattesia. Les couples ont été places en observation pendant 10 semaines. Outre le nombre d'oeufs pondus, on a également noté la longévité des sujets en vue d'en tirer des conclusions quant aux incidences de l'infection par Mattesia sur la fécondité.
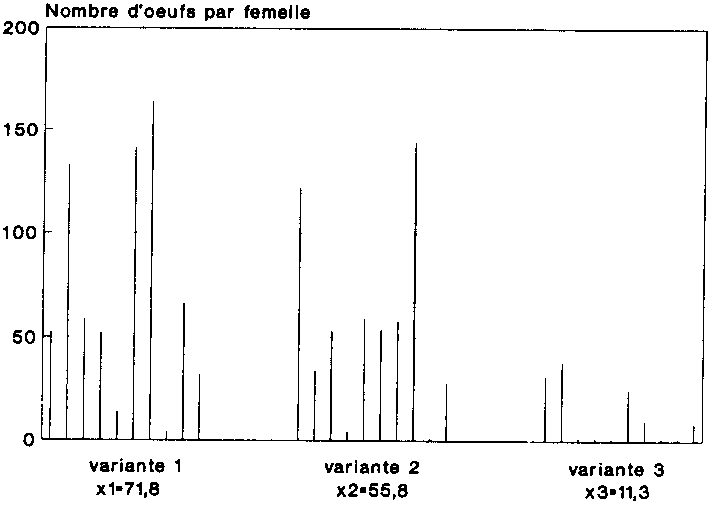
La figure 8 indique la ponte par femelle pour chacune des trois variantes. La production moyenne d'oeufs des femelles infectées était nettement inférieure à celle des femelles saines. Chez les 10 femelles malades, le nombre moyen d'oeufs par individu était de 11,3, la moitié seulement a entre elles ayant produit des oeufs. A l oppose, les femelles saines ont pondu une moyenne de 71,8 oeufs. Les 10 femelles de ce groupe ont toutes été fécondes. Chez les couples comportant un mâle infecte, enfin, chaque femelle a pondu en moyenne 55,6 oeufs, un seul de ces couples étant demeure stérile. On peut établir une corrélation entre ce bilan de ponte, qui présente comme on le Voit des différences tris accentuées selon les variantes, et la longévité des individus.
Le tableau 9 indique la longévité moyenne des miles et des femelles au cours d'une période d'observation maximale de 10 semaines. Les P. truncatus sains ont survécu en moyenne 54 jours, alors que la longévité des femelles infectées était limitée à 31 jours seulement. Quant aux miles infectés, leur viabilité étaient même encore inférieure, puisqu'ils n'ont survécu que 28 jours.
Comme le montrent clairement ces chiffres, la réduction de la ponte chez les couples infestés est a mettre au compte de la longévité fortement écourtée des sujets malades. L'influence de la femelle était en l'occurrence dominante, puisque le nombre d'oeufs pondus dans la variante 2 était, malgré la présence des miles infestés par Mattesia, supérieur à celui enregistre dans le groupe des femelles infestées (cf. fig. 8).
Il ne fait aucun doute que la baisse de reproduction des P. truncatus infectés s'explique par des mutations physiologiques aussi bien qu'histologiques. Au stade avancé de la maladie, le protozoaire envahit progressivement tous les organes de l'organisme hôte, c'est3-dire que l'agent pathogène peut également coloniser les ovaires et les testicules. C'est ce qui explique que dans les couples comportant des mâles infestes, la reproduction ait aussi baissé, même si cette baisse (tait moins accentuée que dans le groupe dont les femelles étaient infestées.
Tabl. 9 : Longévité moyenne des femelles et des mâles de
Prostephanus truncatus à la suite d'une infection par Mattesia
sp. (10 couples sains, 10 couples avec femelles infestées et 10
couples avec miles infestés)
Longévité (en jours) |
|
| Femelles saines | 54 |
| Mâles sains | 54 |
| Femelles infestées | 30,9 |
| Mâles infestés | 27,8 |
Au cours de la période d'observation des couples, on a note chez les sujets infestés par la néogrégarine un symptôme externe. Certains coléoptères' morts au bout de 20 jours déjà, présentaient en effet un abdomen fortement ballonné (cf. fig. 9). On a trouvé dans les préparations contuses de ces animaux une multitude de stades Mattesia. C'est cette reproduction massive de Mattesia sp. qui était donc, de toute évidence, à l'origine du gonflement
Les oeufs des insectes ont été nettoyés à l'eau distillée aussitôt après la ponte pour permettre l'examen des larves écloses, Cette mesure visait a prévenir une infestation des larves fraîchement écloses par d'éventuelles spores de Mattesia adhérant a la coquille de l'oeuf. L'examen au microscope a révélé qu'aucun L1 n'était infesté par la néogrégarine, ce qui permet d'en conclure qu'il n'y avait pas eu, chez le géniteur transmission de l'espèce Mattesia a la descendance.
Mise en évidence de spores dans des excréments de Prostephanus truncatus infestés
Table
des matières - ![]() Précédente
- Suivante
Précédente
- Suivante![]()

{kind=link}
{kind=link}
{kind=link}